Dra. Dianora Navarro Gastroenterólogo pediatra. Coordinador de Postgrado. Unidad de Gastroenterología y Nutrición Pediátrica. Hospital Dr. Miguel Pérez Carreño. IVSS, Caracas-Venezuela.
RESUMEN
La enfermedad celíaca es proceso sistémico inmunomediado provocado por el consumo de gluten en personas genéticamente susceptibles (haplotipo HLA-DQ2/DQ8) con o sin enteropatía. El consumo de gluten es el desencadenante ambiental más importantes. En el artículo se describen los factores ambientales, genético se inmune que contribuyen a la patología y lesión intestinal. Mecanismos sobre inmunidad se han estudiados y actualmente se acepta que en la inmunidad adaptativa existe la participación de linfocitos T CD4+ reactivos al gluten, lo que conduce a una respuesta proinflamatoria dominada por la producción de interferón (IFN-γ) y la interleucina-21 (IL-21). En forma paralela, los péptidos del gluten con una digestión incompleta ejercen un efecto tóxico con transporte transepitelial activo, en este caso la inmunidad innata activa la citotoxicidad epitelial mediada por linfocitos intraepiteliales (LIE). De importancia hay que mencionar que la transglutaminasa 2 (TG2) ejerce funciones como una enzima desamidante, que puede mejorar el efecto inmunoestimulador del gluten, y la otra como autoantígeno diana en la respuesta inmune. La fisiopatología de la EC es compleja, todavía en estudio mucho de los mecanismos involucrados.
Palabras claves: enfermedad celíaca, haplotipo HLA-DQ2/DQ8, linfocitos intraepiteliales, interleucinas.
ABSTRACT
Celiac disease is an immune-mediated systemic process caused by gluten consumption in genetically susceptible people (HLA-DQ2/DQ8 haplotype) with or without enteropathy. Gluten consumption is the most important environmental trigger. The article describes the environmental, genetic and immune factors that contribute to intestinal pathology and injury. Mechanisms of immunity have been studied and it is currently accepted that adaptive immunity involves the participation of gluten-reactive CD4+ T lymphocytes, which leads to a proinflammatory response dominated by the production of interferon (IFN-γ) and interleukin-21 ( IL-21). In parallel, gluten peptides with incomplete digestion exert a toxic effect with active transepithelial transport, in this case innate immunity activates epithelial cytotoxicity mediated by intraepithelial lymphocytes (IEL). It is important to mention that transglutaminase 2 (TG2) exerts functions as a deamidating enzyme, which can improve the immunostimulatory effect of gluten, and the other as a target autoantigen in the immune response. The pathophysiology of CD is complex, many of the mechanisms involved are still being studied.
Key words:celiac disease, HLA-DQ2/DQ8 haplotype, intraepithelial lymphocytes, interleukins.Autor de correspondencia: Dianora Navarro, Gastroenterólogo pediatra. Coordinador de Postgrado. Unidad de Gastroenterología y Nutrición Pediátrica. Hospital Dr. Miguel Pérez Carreño. IVSS, Caracas-Venezuela, correo electrónico: dianora.navarro@hotmail.com
INTRODUCCIÓN
Con el conocimiento de los mecanismos fisiopatológicos de la enfermedad celíaca (EC) su definición ha ido evolucionando en el tiempo, actualmente, se considera como un proceso sistémico inmunomediado provocado por el consumo de gluten y prolaminas relacionadas, en personas genéticamente susceptibles haplotipo del sistema mayor de histocompatibilidad HLA DQ2 / DQ8). Se sabe que puede presentarse con o sin enteropatía. (1)
En estudios de seroprevalencia en la EC con la determinación de anticuerpos positivos antitrasglutaminasa tisular y/o antiendomisio señalan una prevalencia global de 1,4 % y en aquellos con la confirmada por biopsia intestinal fue del 0,7 %. No obstante, los valores de prevalencia son variables dependiendo de la región geográfica para América del Sur 0,4%, en África y América del Norte en 0,5%, con niveles ligeramente más altos en Europa con 0,8% y 0,6% en Asia. (2)
El gluten como componente principal del trigo es el desencadenante ambiental de la enfermedad. El trigo ocupa el tercer lugar como cereal más cultivado en el mundo después del maíz y el arroz. La producción mundial se estima en aproximadamente 600 millones de toneladas métricas para el 2015. (3) El trigo posee propiedades que le permite hacer pan y otros productos de manera industrializada en forma masiva para la nutrición humana y animal dado su viscoelasticidad. Los tipos de trigo más utilizados son el trigo duro o tetraploide (genoma A y B) y el trigo hexaploide (genoma A, B y D). (4)
En el sentido estricto, el gluten define las proteínas de almacenamiento de cereales, no solo del trigo sino también del centeno, cebada y avena. Con la excepción de la avena, que generalmente se consideran seguros, todos estos cereales dan lugar a una respuesta inmune dañina en pacientes predispuestos a la EC. (5) Se ha demostrado con los estudios del genoma humano que en la EC se presenta una adaptación del sistema inmune con relación a la expresión de ciertas variantes de haplotipo HLA-DQ2 y DQ8 que señalan la presentación del antígeno a las células T CD4+ como evento patogénico clave (5).
FISIOPATOLOGIA.
En la patogénesis de la enfermedad celiaca están involucrados factores ambientales, genéticos e inmunológicos. A continuación, se describen estos factores:
1.- Factores ambientales.
Ciertos fragmentos del trigo como la gliadina del gluten estimulan una respuesta inmunitaria tóxica que daña permanentemente el intestino delgado y afecta la absorción de nutrientes. (4, 6) Esta respuesta inmune es distinta a los otros trastornos relacionados con la ingestión de trigo como la sensibilidad al trigo no celíaca y alergia al trigo. Estas dos últimas la respuesta puede surgir del contacto, inhalación o ingestión de trigo, pero además de estar asociada al gluten, responden a otras proteínas de trigo y carbohidratos presentes en este cereal particularmente fermentables, oligosacáridos, disacáridos, monosacáridos y polioles, los denominados FODMAP. (4)
El gluten tiene más gliadinas y glutamina con menos aminoácidos cargados que otros cereales, con un peso molecular natural (PM) de 10 a 30 kDa o más. Algunos péptidos de gliadina son resistentes a la degradación por enzimas del tracto gastrointestinal y permanecen en la luz intestinal como unos péptidos largos, causando estrés y una respuesta inmunológica innata que en individuos genéticamente susceptibles conducirá al desarrollo de EC. (6)
Se ha demostrado que la gliadina puede provocar un aumento inmediato y transitorio de la permeabilidad intestinal, cuyo efecto se ha relacionado con la liberación de zonulina, a nivel paracelular provocando la apertura de las uniones estrechas. (7) Este factor de permeabilidad intestinal desempeña un papel en el desarrollo de la EC, debido a un mayor tráfico de partículas de gluten no digeridas desde la luz intestinal hasta la lámina propia, interviniendo directamente en la etapa inicial de la activación inmunitaria innata, con adición de los cambios en la microbiota intestinal que se han estudiado en la EC. (6,8-9)
También con respecto a la dieta, algunos estudios han señalado que el inicio de la alimentación con consumo de gluten entre los 4 y 7 meses de edad pudiera desencadenarse el desarrollo de la enfermedad a edades tempranas. Aunque existen resultados contradictorios. En especial cuando se asocia a la lactancia humana, ya que en lactantes que la reciben, se reduce el riesgo de aparición de la EC. (4, 9-10)
Adicionalmente, recientes trabajos de investigación mencionan la posibilidad de la activación de la EC con un proceso infeccioso, aunque no se ha explicado el mecanismo verdadero. Se piensa que el aumento de la permeabilidad intestinal observada en las infecciones gastrointestinales permite el paso del gluten a través de la mucosa, o elevan la expresión de la trasglutaminasa tisular (TG2) que puede ocasionar un aumento de péptidos de gluten inmunogénicos. Así mismo, un antígeno extraño como un virus o una bacteria por mimetismo molecular podría iniciar una respuesta contra el gluten. (9)
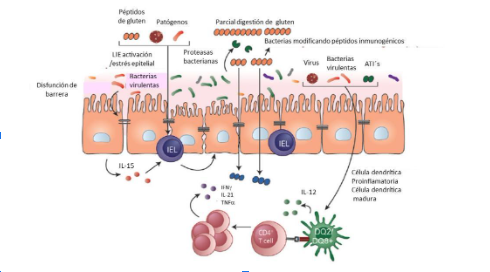
Entre los patógenos investigados y su asociación con EC están el rotavirus, adenovirus, enterovirus y Helicobacter pylori. Se ha estudiado particularmente el rotavirus, y se ha sugerido que la vacunación contra el rotavirus puede proteger contra el desarrollo de EC. (8) Ver Figura 1.
Figura 1. Factores ambientales involucrados con enfermedad celíaca.
2.- Factores genéticos.
El desarrollo de la enfermedad se presenta en personas genéticamente susceptibles en asociación con la presencia del antígeno leucocitario humano (HLA) tipo DQ2 y DQ8. (4)
La tasa de concordancia en EC para los gemelos es cercana a 80% y un 11% para los gemelos dicigóticos. Al igual del riesgo entre los familiares de primer grado, cuya tasa se ha estimado en aproximadamente 10%. (8-9)
Un aporte de los estudios de asociación de todo el genoma ha implicado más de 100 loci de riesgo distintos del HLA en la EC, es decir genes no HLA. Alguno de estos genes distintos codifican moléculas inmunitarias implicadas en la función de las células T y las células B. Se han detectado muchos de los polimorfismos de un solo nucleótido que probablemente median su efecto controlando la expresión genética. (7-8) La relevancia de estos genes adicionales para conferir riesgo genético de EC es bastante limitada, pero pueden conducir al descubrimiento de vías clave potencialmente involucradas en la patogénesis de la enfermedad.
3.- Factores inmunológicos.
Respuesta inmune al gluten.
Se ha comentado en párrafo anterior que el contenido de las proteínas del gluten (prolinas) son resisten a la degradación por las proteasas digestivas, estimulan tanto el sistema adaptativo como innatos del sistema inmunológico desencadenando la respuesta adversa intestinal y lesión. (8,9).
La enzima TG2 se encuentra en el epitelio del borde en cepillo del intestino delgado o en el espacio extracelular de la región subepitelial. La TG2 existe en forma inactivada en condiciones oxidativas normales, y se activa extracelularmente bajo condiciones reductoras creadas por la respuesta inflamatoria durante la EC. (4)
Adicionalmente, la TG, como enzima dependiente de calcio cataliza la modificación postraduccional de proteínas y se libera de las células durante la inflamación. A nivel de la lámina propia de la mucosa, ejerce dos funciones importantes en la EC, la primera como una enzima desamidante, que puede mejorar el efecto inmunoestimulador del gluten, y la otra como autoantígeno diana en la respuesta inmune. (11-12) Anticuerpos específicos contra la TG2, junto con los anticuerpos anti-endomisio que son típicos de la EC, permiten su determinación serológica para el diagnóstico inicial de la enfermedad. (12)
Respuesta inmune innata al gluten.
La inmunidad innata está involucrada en el comienzo de la reacción inmune de la EC, donde las citocinas como la interleucina IL15 y el interferón α (IFNα) pueden estimular la respuesta inmunitaria innata al polarizar las células dendríticas y la función de los linfocitos intraepiteliales (LIE). (7)
Los péptidos de gluten, como el p31–43 de la alfa-gliadina, pueden dañar directamente el epitelio intestinal al activar mecanismos de la inmunidad innata, con producción de IL15 e inducción de apoptosis en los enterocitos, que alteran la función barrera epitelial y el aumento de la permeabilidad. La respuesta inmune se inicia dentro de los 20 minutos posteriores a la exposición del péptido. Se demostró que p31-43 inhibe la subunidad del canal de cloruro (CFTR) y, por lo tanto, causa la activación de linfocitos asesinos naturales (NF-kB), asociado a la inducción de IL-15 y la activación de TG2, con una variedad de consecuencias inflamatorias. (7.13)
La IL-15 induce la proliferación y la activación de LIE T CD8+ que expresan receptores de células NK tipo NKG2D y CD94-NKG2A, cuyos ligandos son las moléculas de estrés MICA/B y HLA-E, respectivamente, expresadas por los enterocitos en situaciones de estrés (8,13). También la IL-15 promueve en los LIE la producción del IFN-γ y la citotoxicidad dependiente de proteínas citolíticas, como perforinas o granzima. La lesión del epitelio depende de los LIE que expresan el receptor de célula T-αβ, que descienden tras la restricción de gluten, mientras que los LIE receptores de célula T γδ NKG2A tendrían una función reguladora, y se mantienen elevados en dieta sin gluten. (13)
Recientemente, se ha conocido que los péptidos de gliadina captados por las células epiteliales mediante endocitosis son capaces de llegar hasta las vesículas paranucleares de estas células (endosomas tardíos y lisosomas); sin embargo, en vez de ser degradados en los lisosomas, el péptido p31–43 se acumula allí por causas aún desconocidas, lo que provoca un microambiente pro-oxidativo que induce la activación de la TG2 y la degradación por la activación del receptor proliferador de peroxisomas (PPARγ), que es una molécula capaz de modular la inflamación intestinal. Este mecanismo podría explicar por qué los pacientes celíacos recaen tras la reintroducción del gluten, incluso antes de que aparezcan signos de inflamación. (13)
Los péptidos de gliadina resultantes de la degradación parcial del gluten en la luz intestinal cruzan la barrera epitelial a través de la vía transepitelial o pasivamente mediante el flujo paracelular accediendo a la lámina propia. La TG2 y estos péptidos de gliadina desamidados son absorbidos por las células presentadoras de antígenos (APC), como las células dendríticas que son proinflamatorias, promueven la activación de las respuestas de las células T CD4+ específicas del gluten en la lámina propia, en relación con una mayor afinidad por las moléculas HLA-DQ2 o DQ8. Debido a su interacción, tanto las células T como las células B se activan produciendo citocinas proinflamatorias, con la destrucción de la mucosa del intestino delgado, luego de la activación de las células T que conduce a la conocida mucosa aplanada o atrófica. (12)
Adicionalmente, la IL-15, como citocina proinflamatoria es producida por células dendríticas, macrófagos, y enterocitos en respuesta al estrés, cuya expresión esta aumentada tanto en el epitelio como en la lámina propia durante la lesión activa. Se presenta en la superficie celular a otras células a través de la cadena IL-15Rα en condiciones de estrés e inflamación. Están involucrados en las respuestas inmunes innatas con un papel central en el reclutamiento se ha sugerido que funciona como una señal de peligro que regula las células T residentes en tejidos y promueve la destrucción de tejidos. (8,12,14)
Respuesta inmune adaptativa.
Las proteínas del gluten en la EC disparan la respuesta inmune adaptativa que involucra las células T y células B. La activación específica de las células inmune comienza con el reconocimiento de los péptidos de gluten desaminado por la TG2. Este proceso de desamidación introduce una carga negativa que tiene una fuerte afinidad por las moléculas del HLA DQ2 y DQ8 presentes en las células dendríticas. Estas moléculas disponen de un bolsillo de unión a péptidos con propiedades únicas para acomodar secuencias peptídicas: DQ2 tiene preferencia por aminoácidos de carga negativa en posiciones centrales (P4, P6 o P7), y en el caso de DQ8, las posiciones son más externas (P1 o P9). (8,15)
Estos péptidos de gluten con la TG2, forman el complejos de enzima-sustrato de TG2-gluten. Se cree que tales complejos impulsan la activación dependiente del gluten de las células B específicas de TG2, lo que conduce a la formación de autoanticuerpos específicos de tTG2, lo que logra explicar colectivamente la respuesta inmune adaptativa en EC. (10) La TG2 induce la sustitución de glutamina de carga positiva por ácido glutámico de carga negativa (glutamina, ácido glutámico o prolina), con formación de otros péptidos el péptido de 33aminoácidos con gran potencial inmunogénico. (15)
Dependiendo de la desamidación postraduccional de los péptidos de gluten por la TG2, se activa la regulación al ascenso de IL-15 y se promueve la respuesta inmunitaria adaptativa de los linfocitos T CD4+ (14). Estos linfocitos T CD4+ reactivos al gluten conduciría una respuesta proinflamatoria dominada por la producción de IFN-γ. Algunas enzimas bacterianas, como la prolilendopeptidasa, inducen la degradación de estos fragmentos e impiden que se formen epítopos T capaces de activar respuestas de la inmunidad adaptativa. (8,15)
La investigación también ha revisado la respuesta adaptativa humoral, la cual tiene una contribución menor en la patogenia de la EC. Los anticuerpos de IgA secretora específicos de gliadina podrían favorecer la translocación o el paso de péptidos de gliadina intactos desde la luz intestinal hasta el interior por una vía en la que intervendría el receptor de la transferrina CD712. No se han identificado linfocitos T específicos de TG2, y falta por explicar aún por qué la ingestión de una proteína de la dieta (gluten) promueve la producción de autoanticuerpos frente a la TG2. Como posible explicación se ha propuesto un modelo en el que los linfocitos T CD4+ reactivos al gluten podrían proporcionar la ayuda necesaria a las células B específicas de TG2 para la síntesis de anticuerpos vía formación de complejos formados por TG2 y péptidos de gliadina. (15)
En un estudio realizado con anticuerpos desaminados, se encontró que los anticuerpos anti-gliadina desaminada son mejores que los anticuerpos contra la gliadina convencionales. Además de anticuerpos específicos de TG2 que se encuentran en el suero como isotipos IgA e IgG y son considerados autoantígenos para la EC. La prueba de IgA específica para TG2 es la práctica clínica más común utilizada en el caso de EC debido a su mayor especificidad y sensibilidad entre todos los demás métodos. (4)
Estas citocinas producidas por células de la inmunidad adaptativa (IFN-γ, IL-21) en interacción con la inmunidad innata (IFN-α, IL-15), podrían determinar el desarrollo de la inflamación y la enteropatía en la EC, además de contribuir a la pérdida de tolerancia al gluten, debido al bloqueo de la vía de señalización del TGFβ, por la IL-15 o por inhibición de la supresión de los linfocitos T efectores por linfocitos T reguladores, a través de la IL-21. (15)
Las citocinas como el IFN-γ es producido por linfocitos T CD4 específicos del gluten y se cree que también las células T intraepiteliales TCRαβCD8 y TCRγδ contribuyan con la producción de esta citocina. Los enterocitos en respuesta a la estimulación de IFN-γ producen las quimiocinas CXCL10 y CXCL11, pueden desempeñar un papel en la migración de linfocitos al epitelio. (13-14)
Lesión intestinal.
Una de las características distintivas en la EC es la lesión intestinal, la cual es empleada como diagnóstico de la enfermedad. Se observa alteración de la estructura de la vellosidad y de la relación vellosidad cripta, e hiperplasia de la cripta. La clasificación de Marsh Oberhuber es utilizada para describir los hallazgos en la mucosa intestinal. (16,17) El enterocito o célula intestinal constituye el 90% del epitelio, en condiciones normales migran desde la cripta hacia la punta de la vellosidad en un periodo de 4-6 días y posteriormente son reemplazadas o renovadas. Se ha estimado que, en humanos, 1011 células epiteliales se eliminan todos los días. Este número aumenta en condiciones inflamatorias. (10, 11) En la EC activa, la tasa de renovación de los enterocitos aumenta a medida que el índice proliferativo se duplica en comparación con el estado saludable, a 1-2 días. (18)
Otras células presentes en la lámina propia, son leucocitos y linfocitos. Los LIE, residen entre las células epiteliales intestinales, aun en condiciones no inflamatorias. En esta enfermedad, el número de LIE aumenta desde un número normal de <25 por cada 100 enterocitos contados en la parte superior de las vellosidades y, su aumento o se considera exclusivo de la EC. Sin embargo, se considera un signo diagnóstico de la enfermedad a nivel de la muestra histológica. (8,14) La exposición al antígeno en la EC provoca una rápida activación in situ de los LIE de células T α/β. Estas células dañan los enterocitos a través de contribuciones de varios mecanismos posibles, incluida la vía del gen A relacionado con la cadena de clase I del complejo mayor de histocompatibilidad NKG2D, la vía del ligando Fas-Fas, el procedimiento de perforina-granzima o las metaloproteinasas de matriz. La IL-15 contribuye notablemente en esta respuesta, aumentar la expresión del ligando del gen A relacionado con la cadena de clase I del complejo mayor de histocompatibilidad en células epiteliales para facilitar la vía de señalización del receptor NKG2D. (8,17)
En humanos, el compartimento LIE está compuesto por células T que expresan receptores de células T (TCR) αβ o γδ, donde las células T αβ pueden ser CD4 o CD8.
Los linfocitos T TCRαβ dominan sobre los linfocitos T CRγδ y entre los linfocitos T TCRαβ, los linfocitos CD8 dominan sobre los linfocitos CD4. (14)
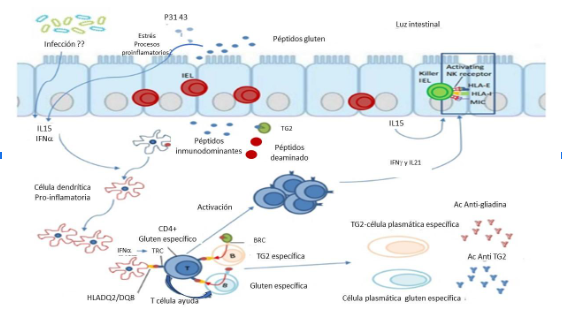
Otras células ´presentes en la lámina propia en la lesión activa de la EC son las células plasmáticas que aumentan en número y son del tipo inmunoglobulina A (IgA), IgM e IgG en el intestino delgado, principalmente en yeyuno, se destaca que hay pocas IgG en el intestino delgado normal y dominan las células plasmáticas que expresan IgA, esta IgA junto con la IgM expresan las inmunoglobulinas de superficie a diferencia de la IgG. Estas moléculas actúan como receptores funcionales de células B (BCR) con co-expresión de CD79ab. Se desconoce el papel de las real de las BCR para las células plasmáticas pero se piensa que sirve de cebado para el antígeno, ya que la célula B reconoce su antígeno (complejo TG2-gliadina o péptidos de gliadina) por BCR, lo internaliza y procesa y lo presenta en el contexto de las moléculas HLA-DQ2 o DQ8 a las células T CD4+ específicas de gliadina (12,14) Además del BCR, las células plasmáticas intestinales tanto en pacientes con EC como en donantes no celíacos expresan bajos niveles de moléculas HLA de clase II, lo que aumenta la posibilidad de que puedan captar el antígeno y presentarlo a las células T CD4. En definitiva, se encontró que las células plasmáticas de la lámina propia son las células presentadoras de péptido de gluten más abundantes en la lesión celíaca.
Figura 2. Factores inmunológicos
COMPLICACIONES.
La EC como proceso inflamatorio crónico afecta el intestino delgado especialmente el yeyuno, que ocasiona una alteración de las vellosidades en el intestino hasta ocasionar una atrofia o aplanamiento, lo que conduce a malabsorción de nutrientes, déficit de vitaminas y minerales. La dieta libre de gluten es el tratamiento de EC, los pacientes que no la reciben desarrollan riesgo de algunas afecciones crónicas y síntomas no gastrointestinales, como anemia, osteoporosis, fatiga, infertilidad, eccema y EC refractaria (ECR), que se asocia con el desarrollo de linfoma. (4)
Pocos pacientes celiacos, generalmente adultos desarrollan una refractariedad a la dieta, y se ha clasificado clínicamente como una ECR. Se distinguen dos tipos, ECR tipo 1, puede deberse a un cambio hacia la autoinmunidad independiente del gluten y ECR tipo 2 es un linfoma intraepitelial de bajo grado, que puede con frecuencia evolucionar hacia enteropatía como el Linfoma de células T, complicación muy rara pero grave. (19)
En lo que se ha denominado “linfogenesis” se descrite que en la ECR tipo 2, estas células malignas, se caracterizan por mutaciones somáticas. En un escenario de cooperación entre las citocinas liberadas por proteínas específicas del gluten, linfocitos CD4+ y el exceso de IL15 en el intestino celíaco, lo que conlleva al ataque del epitelio intestinal, probablemente a través de la activación sostenida de la señal de quinasa Janus, transductor y activador de la vía de transcripción (JAK/STAT) en LIE. Estos linfomas surgen del LIE e inflamación intestinal crónica. (19,20)
CONCLUSIONES
Finalmente, todavía existe los mecanismos que se encuentra bajo investigación. La EC se presenta como proceso fisiopatológico, cada día se identifican más elementos que explican la respuesta inmune.
REFERENCIAS BIBLIOGRÁFICAS
1- Husby S, Koletzko S, Korponay I, Kurppa K, Mearin M, y col. European Society Paediatric Gastroenterology, Hepatology and Nutrition Guidelines for Diagnosing Coeliac Disease 2020. Pediatr Gastroenterol Nutr 2020; 70 (1): 141-156.
2- Singh P, Arora A,Strand T, Leffler D, Catassi C, y col. Global Prevalence of Celiac Disease: Systematic Review and Meta-analysis. Clinical Gastroenterology and Hepatology 2018;16:823–836.
3- Shewry P, Hey S. The contribution of wheat to human diet and health. Food Energy Secur 2015;4(3):178–202.
4- Sharma N, Bhatia S, Chunduri V, Kaur S, Sharma S, y col. Pathogenesis of Celiac Disease and Other Gluten Related Disorders in Wheat and Strategies for Mitigating Them. Front Nutr. 2020; 7:1-26.
5- Rasmus I, Ludvig M. The Immunobiology and Pathogenesis of Celiac Disease. Annu Rev Pathol Mech Dis 2023. 18:47–70.
6- Aboulaghras S, Piancatelli D, Oumhani K, Balahbib A, Bouyahya A, Taghzouti K. Pathophysiology and immunogenetics of celiac disease. Clinic Chimica Acta 2022;528:74-83.
7- Wu X, Qian L, Liu K, Wu J, Shan Z. Gastrointestinal microbiome and gluten in celiac disease. Ann Med 2021; 53(1): 1797–1805.
8- Iversen R, Sollid LM. The Annual Review of Pathology: Mechanisms of Disease. Annu Rev Pathol Mech Dis 2023. 18:47–70.
9- Tye-Din JA, Galipeau HJ, Agardh D Celiac Disease: A Review of Current Concepts in Pathogenesis, Prevention, and Novel Therapies. Front. Pediatr.2018 6:350.
10- Ivarsson A, Hernell O, Stenlund H, Persson LA. Breast-feeding protects against celiac disease. Am J Clin Nutr 2002; 75:914-921
11- Di Sabino A, Vanoli A, Giuffrida P, Luinetti O, Solcia E, y col. The function of tissue transglutaminase in celiac disease. Reviews Autoinmunity 2012; 11(10):746-753.
12- Maglio M and Troncone R. Intestinal Anti-tissue Transglutaminase2 Autoantibodies: Pathogenic and Clinical Implications for Celiac Disease. Front. Nutr. 2020;7:73.
13- Dunne M, Byrne G, Chirdo F, Feighery C. Celiac Disease Pathogenesis: The Uncertainties of a Well-Known Immune Mediated Disorder. Front. Immunol 2020; 11:1374.
14- Gomollón F. Avances en la enfermedad celíaca: un modelo de enfermedad inmunológica. GH CONTINUADA. 2006; 5(6): 257-263
15- Arranz E, Garrot J. Immunology of celiac disease. Gastroenterol Hepatol. 2010;33(9):643–651.
16- Oberhuber G. Histopatología de la enfermedad celíaca. Biomed Pharmacother 2000 54 368–372
17- Dickson B, Streutker C, Chetty R. Coeliac disease: an update for pathologists. J Clin Pathol 2006;59:1008–1016.
18- Ciclitira P, King A, Fraser J. Revisión técnica de AGA sobre la enfermedad celíaca. Asociación Americana de Gastroenterología. Gastroenterología 2001; 120: 1526-1540.
19- Levescot A, Malamut G, Cerf-Bensussan N. Immunopathogenesis and environmental triggers in coeliac disease. Gut 2022;71:2337–2349.
20- Malamut G, Cording S, Cerf-Bensussan N. Recent advances in celiac disease and refractory celiac disease. F1000Research 2019;8:969.